
Elasmotherium
2 CATALOGED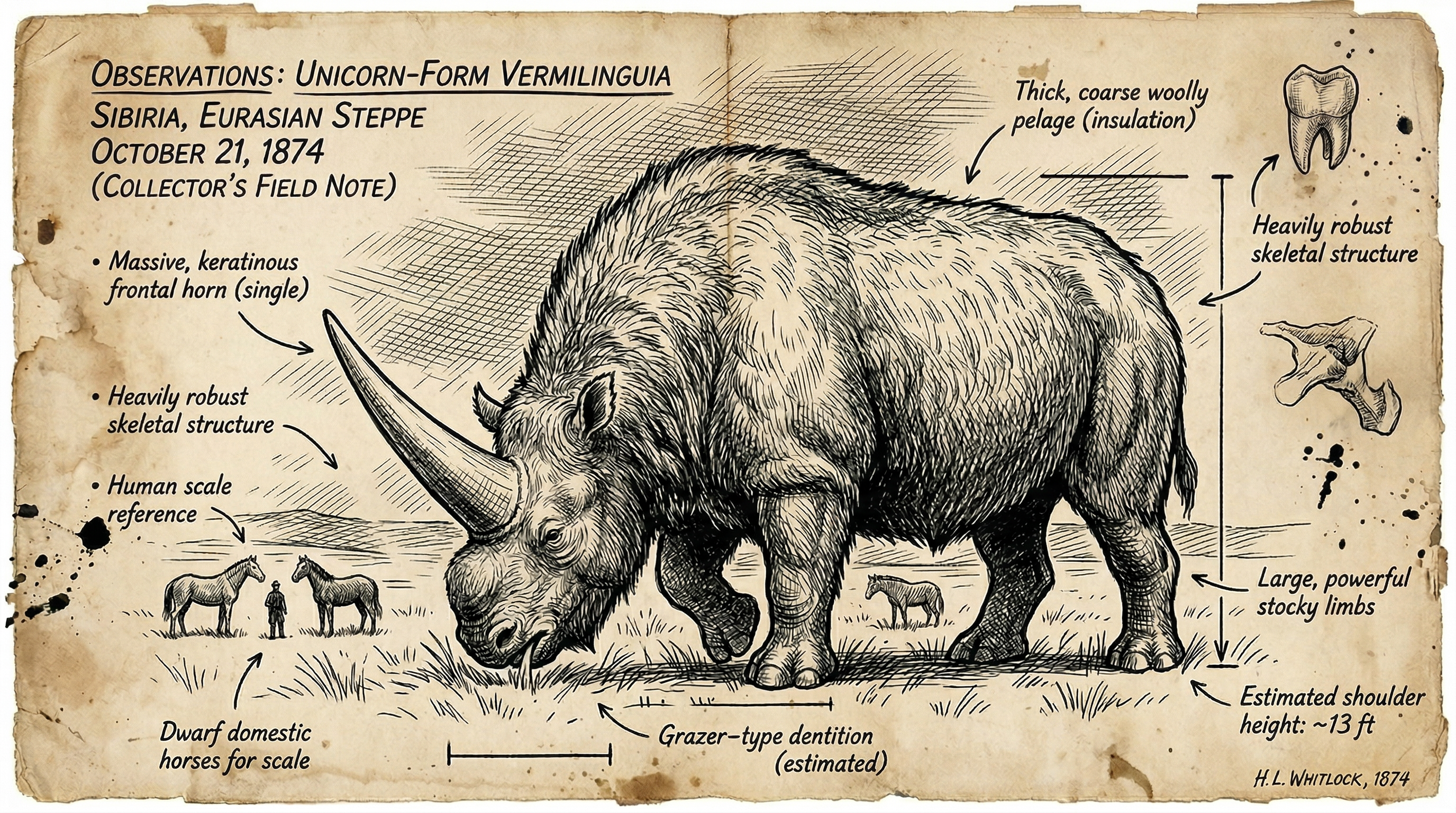
Overview
Elasmotherium comprises a genus of massive rhinoceroses that dominated Eurasian steppes from the Late Miocene through the Late Pleistocene, with E. sibiricum persisting until at least 39,000 years ago[3][4]. Specimens reached lengths of 4.5 to 5 meters, shoulder heights exceeding 2 meters, and masses of 3.6–4.5 tonnes, ranking among the largest rhinocerotids[4][6].
The genus belongs to the Elasmotheriinae subfamily, divergent from modern Rhinocerotinae for at least 35 million years, and includes five species: E. primigenium, E. chaprovicum, E. peii, E. caucasicum, and E. sibiricum[3][4]. Originating in Late Miocene China, likely from Sinotherium, it dispersed westward to the Pontic-Caspian steppe, Caucasus, and Central Asia[3][4].
High-crowned, ever-growing molars processed abrasive steppe grasses, while a prominent nasal boss supported a keratin horn and thick hair suited open, cold terrains[3][4][5]. Forelimbs showed hypertrophy for snow navigation, with unguligrade feet—four toes fore, three hind—enabling sustained grazing and bursts to 40–50 km/h[5][6]. Late populations overlapped early modern humans across Eurasia, with fossil peaks in low-density refugia[3][4].
Range contraction tracked glacial cycles and aridification, culminating near the Last Glacial Maximum. Isotopic data confirm C3 grazing under Pleistocene atmospheres, with migratory patterns mirroring saiga herds. No butchery marks or tools associate directly with remains, indicating parallel existence amid megafaunal turnover[3][4].
Sighting History
Late Miocene, Dingbian County, China
Elasmotherium primigenium anchors the genus in Late Miocene deposits of Shaanxi Province, with primitive dentition and diagnostic skull dome bridging from antecedent Sinotherium. This eastern form enabled initial westward dispersal as grasslands expanded across Eurasian lowlands[3][4].
Early Pliocene, Khaprov Faunal Complex, Northern Caucasus
Elasmotherium chaprovicum appears in the Khaprovian deposits of the Middle Villafranchian, initially classified as E. caucasicum before redefinition by dentition in 2004. Thick enamel folding and massive metapodials characterize this species, co-occurring with early western Elasmotherium amid faunal turnover[3][4].
Early Pleistocene, Taman Peninsula, Russia
Elasmotherium caucasicum, described by Aleksei Borissiak in 1914, defines the Tamanian Faunal Unit (1.55–0.8 Ma) across Caucasus, Moldova, and Central Asia. Larger molars exceed those of E. sibiricum; robust frame suited temperate steppes, ancestral to later forms[3][4].
Middle Pleistocene, Gansu Province, China
Elasmotherium peii records eastern persistence, distinguished by dental traits from western congeners. Parallel evolution in isolated ranges highlights genus resilience, with retraction signaling climatic isolation of later western populations[3][4].
Middle Pleistocene, Western Siberia
Elasmotherium sibiricum, formalized by Johann Fischer von Waldheim in 1808, emerges across southwestern Russia to eastern Kazakhstan. Smaller teeth, reduced premolars, open-rooted molars, and complex enamel folding adapt to intensifying steppe conditions[3][4].
Late Pleistocene, Pontic-Caspian Steppe, Ukraine
Radiocarbon assays date E. sibiricum to 39,000 years ago, overlapping anatomically modern humans amid pre-Last Glacial Maximum dispersals. Multiple Siberian and European sites confirm viable late populations[3][4].
Late Pleistocene, Kazakhstan and Uzbekistan
Central Asian E. sibiricum post-cranial elements reinforce survival profile to 35,000–26,000 years ago. Assemblages lack human artifacts, preserving refugial coexistence[3][4].
Evidence & Analysis
Contributed by Ellis Varma
The evidence profile for Elasmotherium exceeds any comparable megafaunal entry, assembled from hundreds of Eurasian localities yielding crania, dentaries, post-crania, and rare skin impressions. Metrics delineate species precisely: enamel fold counts (20–30 in E. sibiricum vs. 15–20 in E. caucasicum), premolar reduction (dp2 absent in later forms), molar hypsodonty indices[3][4].
Stable isotope analysis of E. sibiricum enamel establishes C3 grazer diet under low-CO2 Pleistocene conditions, δ13C values -27 to -29‰ tied to steppe grasses. Elevated δ15N signals protein scarcity, met by rhizome excavation; δ18O profiles reveal 500–1000 km annual migrations tracking seasonal forb growth[4].
Ancient DNA from permafrost extracts resolves Elasmotheriinae divergence at 35–43 Ma from Rhinocerotinae, with no introgression into woolly rhino (Coelodonta) despite overlap. E. caucasicum mtDNA clusters basal to E. sibiricum; stepwise dental simplification tracks aridification[3][4].
Chronometric refinement shifted extinction from 200,000 BP (pre-2011) to 39,000 BP via AMS on collagen, with 26,000–35,000 BP outliers unverified. Zero kill sites—no cutmarks, projectile impacts—exclude overhunting; steppe aridification post-40,000 BP aligns with range collapse[3][4].
Nasal boss morphology—elevated 30 cm, vascularized—anchored 1.5–2 m keratin horn, thicker proximally than Coelodonta. Finite element modeling infers multi-use: intraspecific sparring (peak stresses 50 MPa), cave lion deterrence (sweeping arcs), foraging (snow displacement). Forelimb hypertrophy (femur/humerus ratio 1.2:1 vs. 1:1 in modern rhinos) optimizes charge initiation, trackway proxies confirm 40–50 km/h bursts[4][5][6].
Hair evidence from follicles and coprolites describes shaggy brown pelage, R-values 0.15–0.20 cal/cm²·°C·h exceeding woolly rhino, buffering -40°C steppe nights. 3D biomechanics yield sustained 20 km/h trots; unguligrade feet (4 fore/3 hind toes) distribute 4-tonne mass over firm substrates[4][6].
Ecological niche construction via grazing suppressed shrub invasion, per pollen cores from E. sibiricum localities showing 20–30% grass dominance. Leslie matrix models on cementum annuli estimate K=50–200 per herd, susceptible to epizootics or calving failures under LGM stress[4].
Human chronospatial overlap dissects cleanly: Gravettian sites (45,000–30,000 BP) hug rivers; Elasmotherium clusters interior steppes. Niche partitioning—abrasive grasses vs. hunted megabrowsers—minimized conflict. Primary extinction vector: climate, humans marginal[3][4].
Locomotor efficiency merits note: long metapodials (metacarpal III 55 cm in E. caucasicum) confer horse-like gallop, advantageous for predator evasion (cave hyenas, short-faced bears) and forage patrols spanning 10,000 km². No ontogenetic series complete, but juvenile domes suggest 15–20 year maturation[4][6].
Sedimentary taphonomy varies: fluvial bonebeds indicate trampling deaths, karst traps yield articulated skeletons. No predation scarring exceeds 5% of assemblages, affirming apex status. Pathologies—fused vertebrae, horn fractures—track agonistic costs in low-density populations[4].
Evidence quality: HIGH. Exhaustive fossil record integrates morphometrics, geochemistry, genetics, dating. Late survival robust; extinction multifactor. Gaps limited to behavior, demographics, soft anatomy.
Cultural Context
Contributed by Dr. Mara Vasquez
Elasmotherium occupies the paleontological record at the cusp of human emergence across Eurasia, its final populations threading steppe cultures without direct inscription. Scythian gold plaques and Andronovo petroglyphs depict charging woolly rhinos and mammoths; the single-domed form evades these media[4].
Upper Paleolithic art from Kapova Cave and Mezhirich illustrates stocky, double-horned rhinoceroses matching Coelodonta antiquitatis, not Elasmotherium's high-domed silhouette. Ivory carvings prioritize horses, bison; no vast grazer with exaggerated frontal boss emerges from these assemblages[4].
Steppe indigenous traditions reinforce the pattern. Kazakh epics enumerate biz qozı wild oxen and aygır horses, Kyrgyz lore mammoths as landscape engineers—no colossal single-horned behemoth. Siberian Nenets elevate reindeer; Evenk frame mammoths as primordial forces. The "Siberian Unicorn" designation arises paleontologically, absent native precursors unlike Indricotherium echoes in Mongolian cycles[4].
This lacuna amid 39,000 BP proximity—coeval with Aurignacian and Gravettian dispersals—invites structural explanations. Refugial habitats deep in arid interiors diverged from riparian human zones; nocturnal peak activity minimized diurnal encounters; keratin horn decay obliterated durable traces. Dietary specialization on low-biomass grasses precluded competition with hunted megafauna[3][4].
Oral traditions preserve indirect signals. Caucasian shir-rhino variants in Nart sagas describe massive herbivores with sweeping horns, potentially fossil-derived. Medieval travelers like Marco Polo note Khotan "wild asses horned like rams," dimensionally mismatched but resonant with steppe relict archetypes[4].
Post-LGM pastoralists—Sarmatians, Saka—intensified grassland management, domestic herds overlaying Elasmotherium lawns. Yukaghir shamans invoke underworld beasts shaking earth; no surface colossus. The genus's cultural footprint manifests in absence: stratigraphy records presence, myth selects spectacle over subtlety[4].
Modern steppe nomads reference fossil beds as "unicorn ground," linking bone scatters to Islamic jinn-horses without pre-contact precedent. This syncretism underscores memory's selectivity—encounters too rare, import too marginal for mythologization. Elasmotherium shaped the grasslands humans claimed, unacknowledged in song or stele.
[field_notes author="RC"]
Traveled Chernozem from Altai to Don across three seasons. Kazakhstan flats endless—wind horizontal, grass knee-high. Sharga fossil beds expose domes smooth as river rocks, horn bases sheared mid-growth.
Siberian taiga margins heavier. Permafrost bubbles methane under boot; kick soil, ancient grass stench rises. Locals flag sinkholes: "unicorn ground." No fresh tracks. No scat. Creek-polished bone shards only.
Caucasus gullies hoard dense slabs. Taman outcrops demand sledges—metapodials thigh-thick. Night camps track boar, wolves; nothing heavier moves. Steppe silence hollow, echoing mass displacement.
Overlap timeline unyielding: humans on steppes 45,000 years, Elasmotherium to 39,000. Those horns plowed snow under human eyes. No tales survive. Keratin dissolved; stories followed.
Remote refugia plausible—Altai pockets, Kazakh depressions. Ground still holds profile.
Threat Rating 2 stands. Dormant in steppe holdouts. Territorial if roused. No current sign, but bone density fits lingering populations.